Hofmeister's Rules - Dynamical System Model - Results
|
||
A Mathematical ModelWe present the dynamical system model of phyllotaxis which has been the object of our research. It is a mathematical derivation of a model by the physicists Douady and Couder, which itself is based on observations by the 19th Century botanist Hofmeister. This model reproduces all spiral patterns exhibited by plants and explains why Fibonacci phyllotaxis is predominant. The applet Dynamical Model gives a simulation of this process.
|
Hofmeister's Rules |
||||
In 1868 the botanist Wilhelm Hofmeister published his microscopic study of meristems. He proposed that in spiral phyllotaxis, botanical units form one by one with the newest in the least crowded spot. Following Douady and Couder (1996, Part I), we make these rules a bit more precise:
|
||||
 |
||||
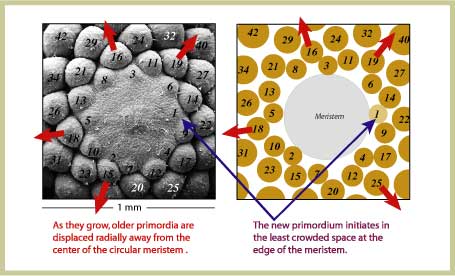
| Compare a scanning electron micrograph of a Norway
spruce (Picea abies) shoot apical
meristem with a computer generated arrangement of primordia. The
computer program implemented the mathematical model based on Hofmeister's
rules and converged naturally to this configuration. Primordia are
numbered according to their age - the higher the number, the older
the primordium. Notice the remarkable similarity between plant and
model.
|
 |
||||
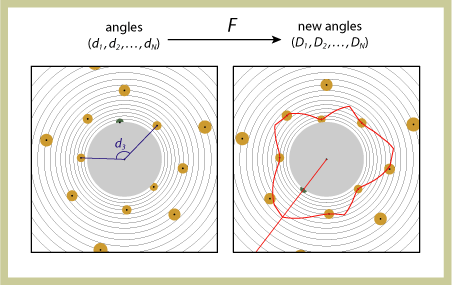
| Under the function F, points of a configuration move out radially to the next circle. The outermost point is eliminated and a new point (green) is generated on the innermost circle, where the distance to the closest point is maximum. The "distance-to-closest" function along the inner circle is represented as a graph (red).
|
Results |
||||
Stationary points in Dynamical Systems The first feature that one usually studies in a dynamical system is its fixed, or stationary points. These are points in the state space which do not change under the map F. For example, the dynamical system given by squaring real numbers, f(x)=x2 has fixed points 0 and 1, since f(0)=0 and f(1)=1. An important characteristic of fixed points is their stability. In the example of the squaring map, 0 is stable: if you start with a point close to 0 (e.g. 0.01), repeated squaring will only get you closer to 0. On the other hand, the fixed point 1 is unstable: repeated squaring of numbers greater than 1 will indefinitely increase these numbers, whereas repeated squaring of numbers smaller than 1 will decrease to 0; in either case, you will get farther and farther from 1. As a rule of thumb, if the dynamical system considered models a natural phenomenon, stable fixed points are significant in the sense that they can be perceived in experiments, as their stability allows for a margin of error in measurements. Stationary configurations in the Phyllotaxis Model In the model F described above, we have the following results (see Hotton (1999) and Atela, Golé & Hotton (2002)):
|
||||
|
||||
| A Scenario for Fibonacci Phyllotaxis These three results put together provide -given Hofmeister's rules- an explanation for the occurrence of spiral phyllotaxis in nature, as well as for the predominance of Fibonacci phyllotaxis in spiral phyllotaxis. Indeed, the expansion parameter G is known to decrease as plants grow, especially at the flowering stage. With large G the pattern has no choice but to converge toward a distichous phyllotaxis. As G decreases, and primordia are formed (i.e.. while the dynamical system is iterated), the stability of stationary configurations forces the nearby configurations to hover along one of the paths of fixed points which starts at distichous, that is, one of the two Fibonacci paths. As G decreases (moving away from the center in the picture), parastichy numbers increase along the Fibonacci sequence, and the divergence angle tend the Golden angle (upper path) or its complement (lower path). Such a scenario is numerically - and visually - verified in the Dynamical Model applet. |
||||